Aujourd’hui, parlons des Cervidae (enfin), famille de mammifères ruminants à bois plus ou moins grands, dont le renne fait partie.
Les Cervidae font partie de l’ordre des Cetartiodactyla. On peut encore voir ordre des Artiodactyla
sur Wikipedia mais ce taxon est aujourd’hui désuet : comme il est maintenant prouvé à partir d’analyses génétiques que les Cetacea (mammifères marins autres que les Pinnipèdes) sont plus proches des Hippopotamidae (les hippopotames) – qui sont des Artiodactyla – que n’importe quel autre taxon, on parle donc de Cetartiodactyla pour y englober les Cetacea (je me rends compte que je parlais d‘Artiodactyla dans mes posts sur le bœuf et la chèvre naine l’été dernier, j’ai honte). Dans l’édition de 2001 de la Classification phylogénétique du vivant
de Lecointre et Le Guyader, on parle bien de Cetartiodactyla (avec Ruminantia, Tylopoda, Suiformes, Hippopothamidae et Cetacea). Artiodactyla est donc un taxon paraphylétique (groupe qui comprend un ancêtre commun mais pas tous ces descendants).
Pour rappel, un taxon est une unité de la classification des espèces vivantes, comme un ordre, une famille, etc. Les arbres phylogénétiques permettent de classifier le monde du vivant et je vais vous montrer des arbres qui respectent le système des clades. Un clade est un taxon monophylétique, c’est-à-dire que les espèces qu’il rassemble descendent d’un même ancêtre commun (une espèce dont ils ont évolué) qui leur est propre. Les nœuds et la construction de l’arbre indiquent alors les degrés de parenté entre chaque clade, c’est-à-dire leurs relations de “cousinage”. Cela permet de classer les espèces vivantes. On peut faire apparaître les informations que l’on souhaite dans un arbre phylogénétique : on peut détailler chaque clade jusqu’aux espèces, s’arrêter aux familles comme ici, ou aux genres, etc.
Il existe d’autres systèmes et certains permettent de prendre en compte les taxons polyphylétiques. Ceux-ci sont des taxons qui regroupent des espèces dont leurs ressemblances sont dues à une convergence évolutive et non à un héritage provenant d’un ancêtre commun. Par exemple tous les mammifères marins forment un taxon polyphylétique puisqu’ils ont convergé vers l’adaptation au milieu marin, mais n’ont pas tous le même ancêtre commun (les baleines sont très éloignés des otaries par exemple) et n’ont donc pas tous évolués de la même façon pour avoir cette caractéristique.
Et donc tout ça pour dire que j’utilise le système des clades (la cladistique) car il permet de mettre en avant l’évolution des espèces et des différents taxons.
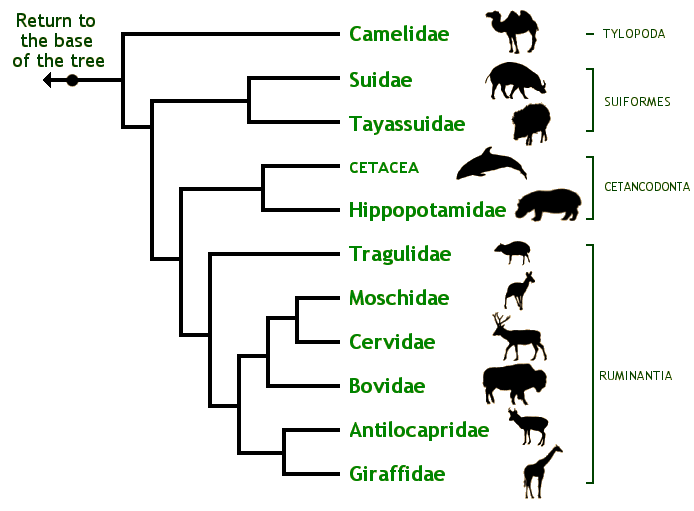
Arbre phylogénétique des Cetartiodactyla © Brent Huffman, www.ultimateungulate.com
Ci-dessus, vous pouvez voir un arbre phylogénétique de l’ordre des Cetartiodactyla. Il montre aussi à droite, le sous-ordre des Ruminantia, et c’est justement dans ce sous-ordre que sont classés les Cervidae.
Chaque clade a des caractères dérivés propres (je pense que le nom est assez explicite). Toujours d’après Lecointre et Le Guyader, pour les Cetarciodactyla, il s’agit par exemple d’un nombre de doigts pair sur les pattes. Le corps repose donc sur les doigts III et IV, puisque leurs ancêtres ont perdu les I, II et V au cours de l’évolution. Pour ce caractère, on les oppose à l’ordre des Perissodactyla (comme les Rhinocerotidae ou les Equidae) qui ont un nombre impair de doigts. Les Cetartiodactyla ont d’autres caractères dérivés propres, comme le talus avec une double poulie ou certaines particularités génétiques.
Au niveau des Ruminantia, il y a par exemple le fait que l’estomac comporte quatre poches (l’humain n’en a qu’une), pour pouvoir ruminer. Dans les Ruminantia, les Cervidae font partie de l’infra-ordre des Pecora, dont le caractère principal est la présence d’excroissances sur le crâne (parfois chez un seul sexe, parfois chez les deux). Cet infra-ordre comprend également les Antilocapridae, les Bovidae (qui portent des cornes avec une enveloppe de kératine) les Giraffoidea (qui portent des ossicônes) et les Moschidae (qui portent des canines transformées en défenses). Les Tragulina (chevrotains), sont l’autre infra-ordre des Ruminantia, avec une famille unique, les Tragulidae, qui n’ont donc pas d’excroissances sur le crâne (ça devient compliqué, mais je vous laisse vous référer à la figure ci-dessus).
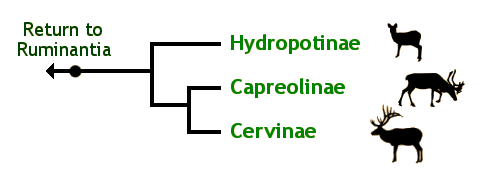
Arbre phylogénétique des Cervidae © Brent Huffman, www.ultimateungulate.com
Ci-dessus, vous pouvez voir un arbre phylogénétique de la famille des Cervidae, qui se limite aux trois sous-familles. Leur caractère dérivé propre est la présence de bois, organes osseux que les mâles portent sur le crâne et qui tombent chaque année, à l’exception de l’hydropote (Hydropotes inermis), qui n’en a pas (caractère que ses ancêtres plus proches ont dû perdre au cours de l’évolution). Une quarantaine d’espèces font partie de cette famille. Les trois sous-familles, comme la figure l’indique sont les Capreolinae, les Cervinae et les Hydropotinae.

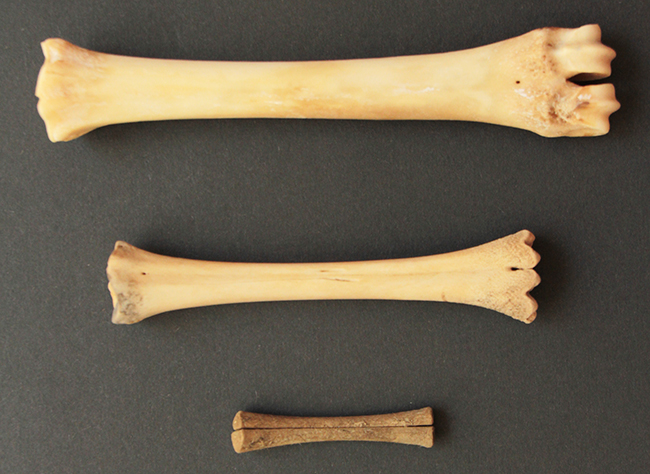
Différents bois de Chevreuil européen (Capreolus capreolus).
Chaque année les bois poussent sur le crâne, un peu plus grands à chaque fois. Sa croissance est stimulée par la production de testostérone et rendue possible grâce au velours. Le velours est une mince couche de peau couverte de poils très doux et très irriguée, qui permet de nourrir le bois durant sa croissance. Lorsque les bois atteignent leur taille définitive, le velours tombe, sanguinolent, pour révéler l’os des bois, alors mort puisque plus qu’il n’est plus irrigué.

Bois en velours de chevreuil européen (Merci Emeline !).
La taille et la bonne formation des bois sont liées à l’âge, l’état de santé et de stress de l’animal et la disponibilité en nourriture. Parfois on peut voir des bois difformes, hypotrophiés, ou monstrueux et hypertrophiés, suite à de grandes périodes de stress ou de détresse de l’animal, ou à des problèmes hormonaux.
Il y a un chapitre très intéressant sur l’évolution, la sélection sexuelle et la fonction des bois dans Evolution de De Panafieu. Vous pouvez aussi trouver des articles de Stephen Jay Gould sur la taille des bois du mégalocéros et la cause de l’extinction de cette espèce.
de De Panafieu. Vous pouvez aussi trouver des articles de Stephen Jay Gould sur la taille des bois du mégalocéros et la cause de l’extinction de cette espèce.
Pour revenir deux minutes sur les autres Pecora, qui portent des cornes et des ossicônes, je voulais insister rapidement sur les différences entre ces appendices. Les bois sont des organes osseux très denses, qui poussent, meurent et tombent chaque année. Les cornes sont des bases osseuses très poreuses que l’animal porte toute sa vie, que l’on appelle également cheville osseuse et qui sont recouvertes d’une couche de kératine (protéine qui constitue en grande partie les cheveux, poils, ongles). Les ossicônes des Giraffidae sont également des bases osseuses, mais recouvertes en permanences de peau et de poils.
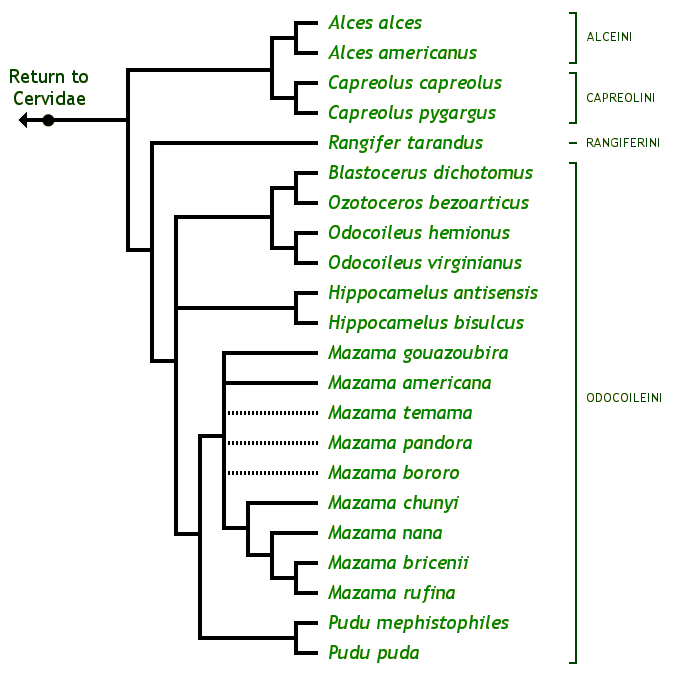
Arbre phylogénétique des Capreolinae © Brent Huffman, www.ultimateungulate.com
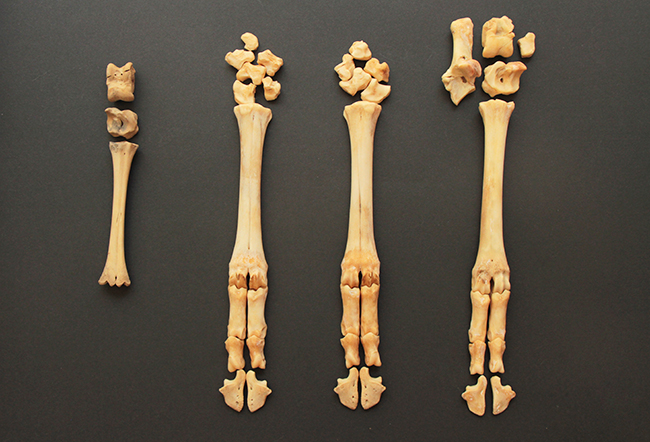
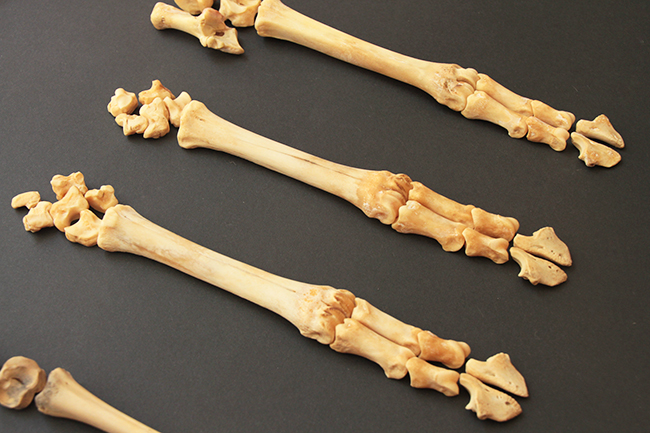
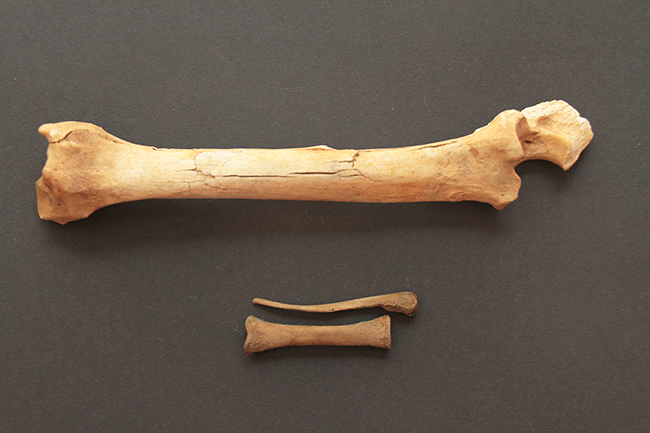
Ci-dessus, vous pouvez voir un arbre phylogénétique de la sous-famille des Capreolinae avec toutes ses espèces. Ce clade est caractérisé par la présence de métacarpes II et V vestigiaux (ils sont très réduits, et ne peuvent conserver leur utilité première, ce sont donc des vestiges), présents contre la partie distale des métacarpes III et IV (qui ont d’ailleurs fusionnés en un seul os). Les métacarpes sont les os qui font la jonction entre poignet et doigts, et comme chez les Cervidae ne restent que les III et IV. On a donc deux mini-doigts avec des mini-sabots présents sur la partie basse des métacarpes III et IV, juste au-dessus des doigts.
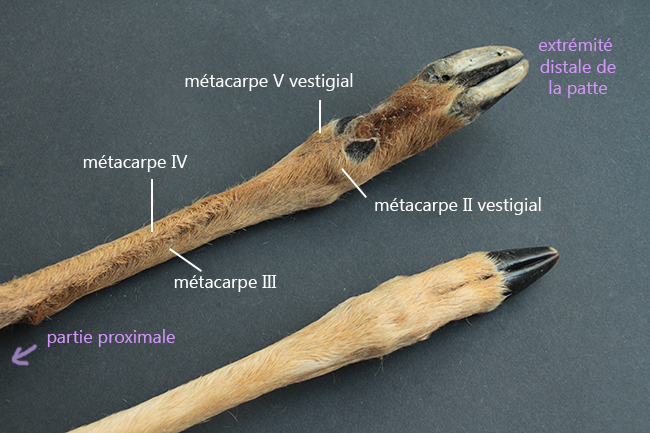
Voici une patte avant droite de chevreuil européen (Capreolus capreolus – merci Émeline bis). Cette espèce fait partie des Capreolinae car ses métacarpes vestigiaux se situent contre la partie distale des autres métacarpes.
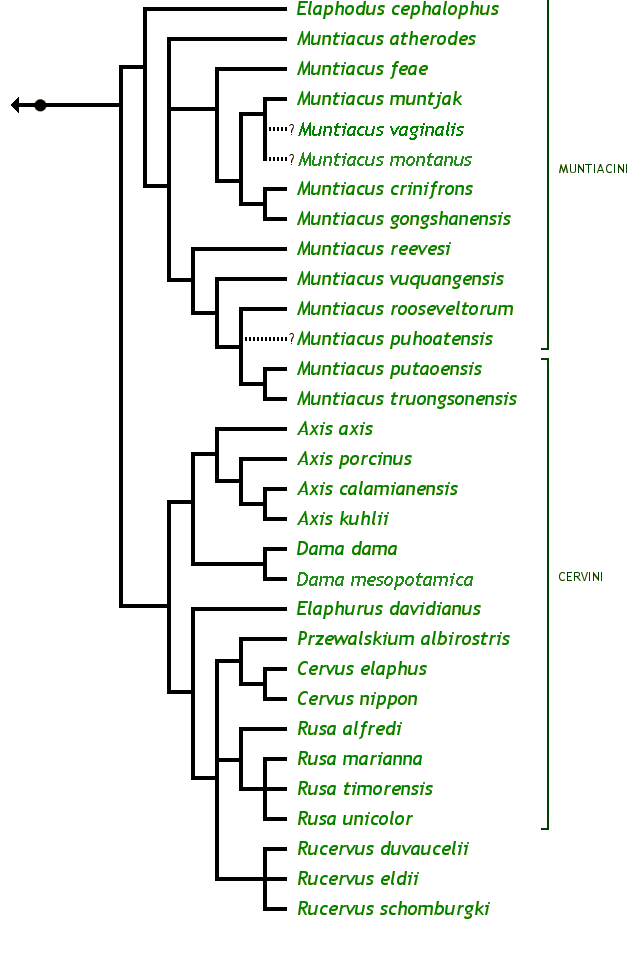
Arbre phylogénétique des Cervinae © Brent Huffman, www.ultimateungulate.com
Ci-dessus, vous pouvez voir un arbre phylogénétique de la sous-famille des Cervinae avec toutes ses espèces. Ce clade est caractérisé par la présence de métacarpes II et V vestigiaux, présents contre la partie proximale cette fois des métacarpes III et IV. On a donc deux mini-doigts avec des mini-sabots présents sur la partie haute des métacarpes III et IV, juste au-dessous du poignet.

Arbre phylogénétique des Hydropotinae © Brent Huffman, www.ultimateungulate.com
La sous-famille des Hydropotinae n’est représentée que par une espèce, l’hydropote (Hydropotes inermis). Comme je l’ai déjà évoquée, sa particularité est de ne pas avoir de bois. J’imagine qu’il s’agit d’un caractère que ses ancêtres plus proches ont dû perdre au cours de l’évolution.
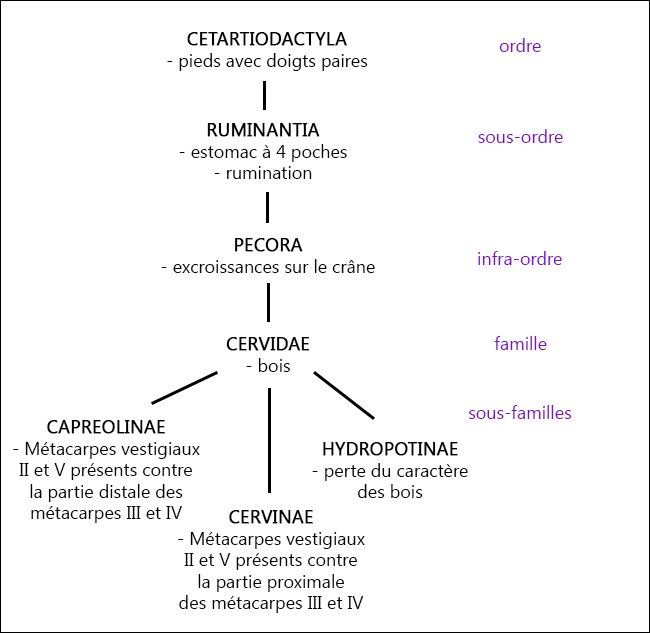
Je vous ai fait un bout d’arbre récapitulatif de l’ordre des Cetartiodactyla jusqu’aux sous-familles de Cervidae, avec les caractères dérivés propres et/ou principales caractéristiques (je ne suis pas sûre que tous soient des caractères dérivés propres, notamment au niveau des sous-familles).
On peut trouver les différentes espèces de Cervidae dans des climats et biotopes très différents. Ces espèces ont une organisation sociale, avec une hiérarchie corrélée à la taille des animaux. Vous pouvez retrouver beaucoup d’informations générales sur cette famille, puis par espèces sur le site animaldiversity (morphologie, répartition, comportement, leur rôle dans leurs écosystèmes, etc).
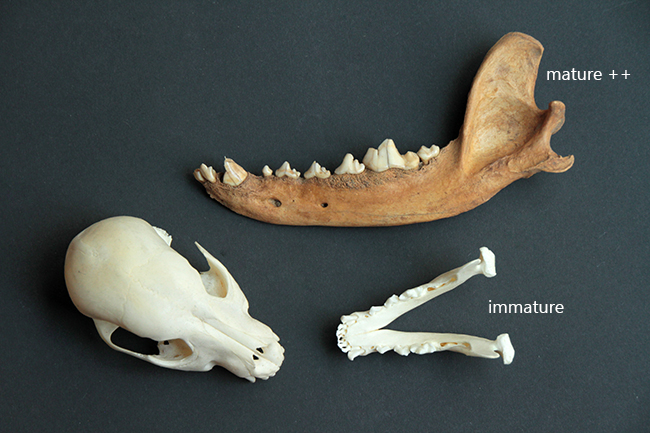
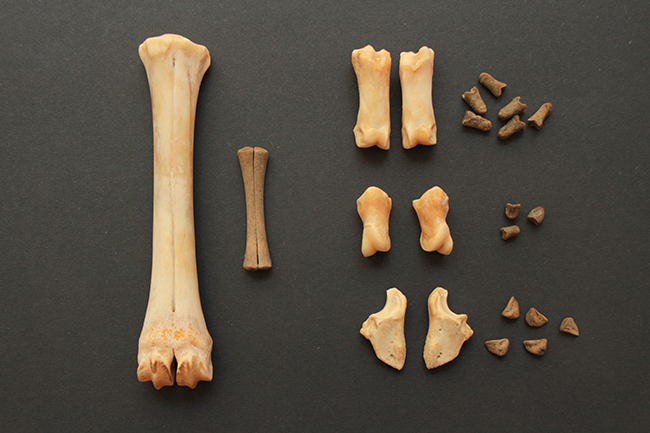
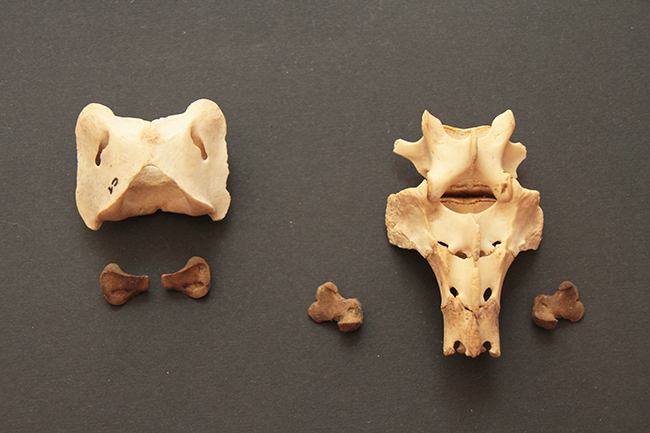
Voici deux crânes de mâles que j’ai à vous montrer, un de chevreuil européen (Capreolus capreolus) à gauche (merci Nicolas), malheureusement partiel, et un d’hydropote (Hydropotes inermis) justement.
Le chevreuil européen est un Cervidae de taille moyenne (65-75 cm de hauteur au garrot, 16-32 kg) qui se répartit dans quasiment toute l’Europe. C’est un Cervidae « typique », sans canine supérieur et avec des bois, uniquement chez le mâle. Ceux-ci font jusqu’à 26 cm de long, ils sont petits, rectilignes, avec faible nombre de pointes.
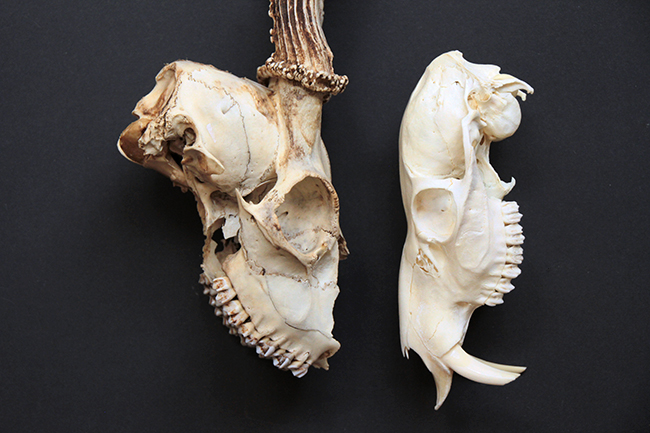
L’hydropote est une petite espèce (45-55 cm au garrot, 9-14 kg) qu’on trouve en Chine, Corée du Sud et du Nord. Il existe également une petite population en Angleterre (le mien vient de là) et une dans le Limousin. Au niveau du crâne, ses particularités sont qu’il n’a pas de bois (c’est donc la seule espèce de Cervidae sans), il a des canines supérieures, et celles-ci sont transformées en défenses chez les mâles. Elles ont le même rôle que les bois, à savoir qu’il s’agit d’un caractère sexuel secondaire, puisque les mâles se battent et se défendent avec durant la période où ils rentrent en compétition pour la reproduction.
En France, mise à part ces deux espèces, on trouve également le daim européen (Dama dama) qui s’y fait très discret (pour les lyonnais il y a une grande population au Parc de la Tête d’Or), le cerf élaphe (Cervus elaphus) et le cerf sika (Cervus nippon, introduit en petites populations à quelques endroits à la fin du XIXe siècle).
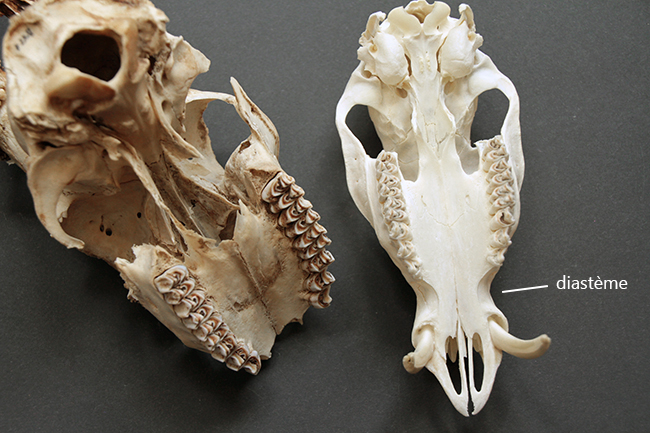
Sur les crânes de Cervidae, on peut voir des adaptions morphologiques liées à leur régime alimentaire herbivore. Ils n’ont pas d’incisives ou canines supérieures et les végétaux sont alors broyés contre leur palais, recouvert d’un tissu très dur. Ils ont un diastème (espace entre les dents labiales – incisives et canines – et jugales – prémolaires et molaires). Leurs prémolaires et molaires sont hypsodontes (à croissance continue, avec une couronne haute) et sélénodontes (les cuspides – ce qui donne le relief à la surface de la couronne – sont en forme de croissant de lune). Leur formule dentaire est 0/3 I 0-1/1 C 3/3 P 3/3 M.
Le rostre est très long, ce qui permet une meilleure attache aux muscles masseter (muscles de la mastication). Leur crâne n’a pas de crête sagittale (celle dont je vous parle tout le temps avec les Carnivora, qui est la fusion des os pariétaux).
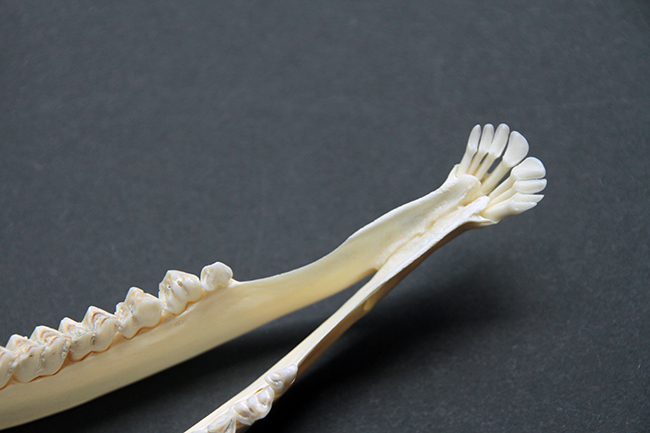
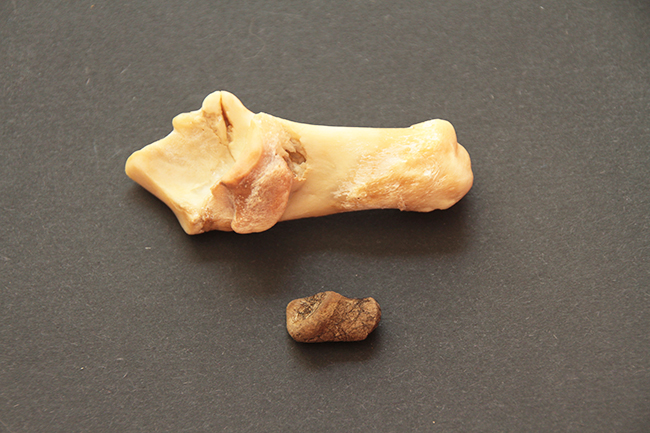
Les fines mandibules de l’hydropote avec un long diastème entre dents labiales et jugales.
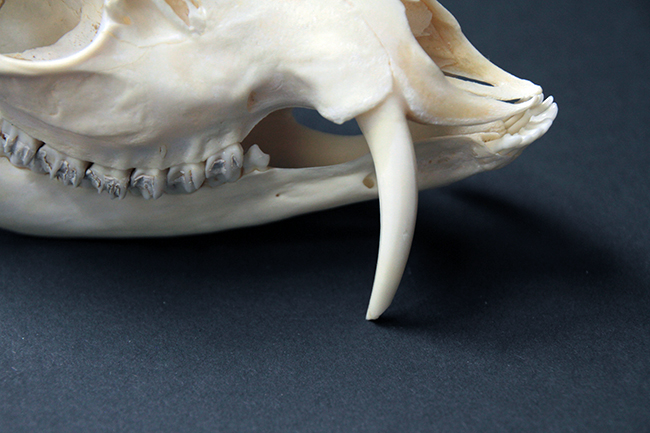
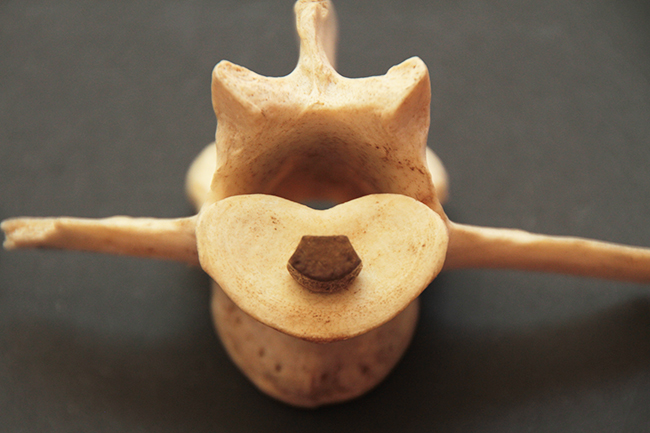
La défense du mâle hydropote, qui prend racine dans l’os maxillaire.

Le bois de chevreuil.

Bois de chevreuil en velours, et bois mort.

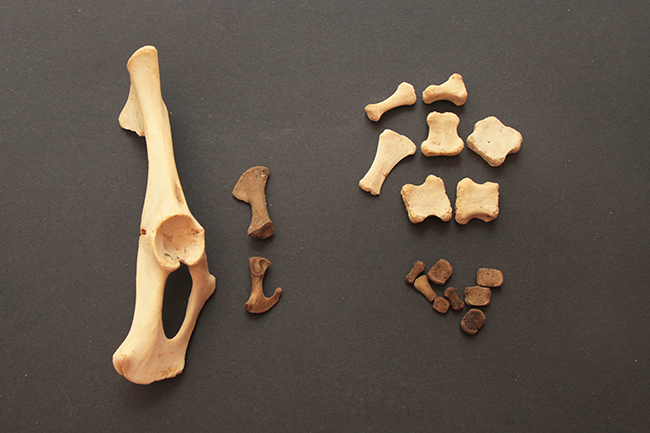
Bois de cerf élaphe (Cervus elaphus).
Le cerf élaphe est le plus grand Cervidae que l’on trouve en France (120-150 cm au garrot, 80-300 kg). C’est une espèce chez qui le dimorphisme sexuel est assez marqué, au niveau de la corpulence, de l’encolure et du pelage (sans parler des bois). Ce bois fait seulement 55 cm (et 724g) et compte cinq pointes, mais les bois de cerf peuvent être beaucoup plus imposants et avoir plus d’une douzaine de pointes.


Bois de daim européen (Dama dama).
Le daim européen est une grande espèce (85-110 cm au garrot, 25-80 kg) dont les bois sont imposants et palmés (avec deux pointes avant la palme). Je ne les ai pas sous la main donc je vous ai mis d’anciennes photos et je n’ai ni la taille (de souvenirs, chaque bois fait entre 70 et 80 cm de long, avec une envergure entre les deux de plus d’un mètre) ni le poids, mais le tout fait plusieurs kilogrammes.

Bois de renne (Rangifer tarandus).
Nous n’en avons pas en France, mais voici un bois de renne (merci les puces !). Il est facilement différenciable du bois de cerf, car il ne part pas de façon rectiligne avec les pointes plutôt vers l’arrière ; mais est plutôt arrondi, beaucoup plus fin, avec des pointes qui poussent en deux bouquets, l’un à l’avant et l’autre à l’arrière. Les bois du mâle peuvent faire 120 cm de long et ceux de la femelle 50 cm. Et oui, le renne est la seule espèce de Cervidae dont la femelle est également pourvue de bois. Les miens sont petits (37cm) et ont une forme assez anarchique.
Le renne est une grande espèce (105-120 cm au garrot, 40-150 kg) qu’on retrouve en Russie, Scandinavie et au nord de l’Amérique du Nord (et en Écosse sous sa forme domestiquée). C’est une espèce domestiquée depuis environ 5000 ans, dont les troupeaux domestiques sont une importante ressource (viande, os, peau, graisse, transport, etc) pour les humains vivants dans les régions les plus froides du monde. Il possède des sabots très imposants, en forme de croissant de lune, adaptés au déplacement dans la neige et la boue (et qui lui permettent aussi de gratter pour trouver sa nourriture). Il a des poils longs et creux qui ne le gênent pas durant la natation, et sa queue est très courte (aucune raison de perdre de l’énergie à irriguer une longue queue à l’utilité réduite quand il fait aussi froid).
Il existe plein d’autres espèces de Cervidae, certaines avec des particularités étonnantes, comme l’élan (Alces alces) qui a les bois les plus imposants ; le muntjac de Reeve (Muntiacus reevesi), petit Cervidae d’Asie du Sud-Est avec de petits bois et de grandes canines ; le pudu (Pudu puda), d’Argentine et du Chili avec de minuscules bois réduits à l’état de vestige, qui fait la taille d’un lièvre, …
Je voulais rapidement reparler des Moschidae, famille que j’ai mentionné en parlant des Ruminantia et des Pecora. C’est le clade le plus proche des Cervidae, et il était auparavant considéré comme une sous-famille de ces derniers. Ce sont les cerfs porte-musc, et dont les ancêtres n’ont jamais vu apparaitre le caractère des bois, ils sont donc plus “primitifs” que les Cervidae. Ils ont par contre une vésicule biliaire (caractère que les Cervidae ont perdu), des canines transformées en défenses et des glandes abdominales sécrétant du musc chez les mâles. Au niveau du crâne, ils ressemblent donc beaucoup à l’hydropote, même s’il a tout de même quelques différences (le crâne de l’hydropote a des os frontaux plus concaves, des incisifs de forme différente, etc).
Sources :
Classification phylogénétique du vivant, de Lecointre et Le Guyader
Evolution, de De Panafieu
Guide des mammifères d’Europe, d’Afrique du Nord et du Moyen-Orient , ed. Delachaux et Niestlé
, ed. Delachaux et Niestlé
Animal Skulls: A Guide to North American Species , de Elbroch
, de Elbroch
http://animaldiversity.org/accounts/Cervidae/
http://tolweb.org
http://animaldiversity.org/collections/mammal_anatomy/horns_and_antlers/
http://www.lejdc.fr/nievre/actualite/2014/04/13/le-cerf-aux-bois-de-velours_1964519.html
http://www.ultimateungulate.com/
http://bioweb.uwlax.edu/bio203/s2009/nelson_lau4/index.htm